![第4回 真核生物の誕生1 1.真核生物は古細菌から生まれた [分子生物学講義Web中継~生物の多様性と進化の驚異]](images/banner03.gif)
- [SHARE]
- ツイート
")

第4回 真核生物の誕生1
- 真核生物は酸素濃度が低ければ生まれなかった!
- クレン古細菌と真核生物は1つのグループ!?
- ・・・など,驚きの視点が満載.
1.真核生物は古細菌から生まれた
真核生物はいつ誕生したか
真核生物は,真核生物らしい大型の化石という証拠から,21億年前には誕生していたと考えられます.縞状鉄鉱床の中から発見された幅0.5mmくらいで長さ数センチ程のリボン状の化石が,グリパニアと名付けられています.形から多細胞の藻類との想像をされていますが,詳しいことは不明のようです.
真核細胞の大型化と酸素濃度
2009年のアメリカ科学アカデミー紀要に,地球の歴史をみると,酸素濃度の上昇がある段階を超えたところで,生物の大きさに飛躍的な巨大化が起きている,という論文があります.約27億年前に誕生したシアノバクテリアによって遊離酸素の濃度が上昇し,約20億年前に現在の濃度の1%(パスツール点)を超える辺りで,細菌に比べて108~109の桁で体積が大きい単細胞真核生物が誕生した,というわけです.酸素は拡散によって細胞内に入るわけで,細胞のサイズが大きくなれば内部の酸素濃度は低くなる.好気的代謝を獲得・維持し,大型の真核生物が出現するためには,パスツール点を超える酸素濃度が必要だったと考えられます.
小型の真核細胞の可能性は…
真核生物らしい大型細胞が誕生する前に,小型の真核生物が誕生していた可能性については,微化石の形態からでは原核細胞と小型真核細胞の区別が十分にはつきません.化学化石の証拠としてはオーストラリアのピルバラで,27億年前の頁岩からステラン(ステロイド類)が検出されており,これは真核生物の細胞膜に特有の成分であることから,真核生物の誕生はそこまで遡る可能性も考えられます.取りあえずここでは,大型細胞出現がほぼ確実と思われている21億年前に真核生物が誕生したことにして話を進めます.
基本は古細菌
真核生物の細胞になったもとは,遺伝子の解析から古細菌とされます.古細菌が真核生物のヒストンH3やH4と類似したタンパク質をもっていて,DNAと複合体を形成してクロマチンのような構造を作っているとか,イントロンをもつ遺伝子の構造や,DNA複製機構にかかわるタンパク質や酵素のさまざまな性質,mRNAの転写のしくみや,それを使ったタンパク質合成系のしくみなど,生物としての基本的な機構に両者の共通性が高く,真正細菌とは離れていることが支持されます.
細胞壁をもたない古細菌の可能性
真核生物の細胞は細菌のような細胞壁をもっていないので,はじめから堅い細胞壁をもたない古細菌に由来するか,あるいは初期の過程で細胞壁を失ったものである可能性があります.
現在生きている古細菌のなかで細胞壁をもっていないものを探すと,サーモプラズマがそうです.通常の古細菌は1μmかそれ以下のサイズですが,サーモプラズマは,成長してDNAを増やしても細胞分裂せずに大きくなる場合や,細胞壁がないために細胞同士が容易に融合し,20μmものサイズになるといわれます.これは真核生物の細胞と同等レベルの大きさです.サーモプラズマには,原始的ではあるけれども,真核生物のオルガネラのような細胞内膜構造をもつものがあります.また,サーモプラズマを含めて古細菌のなかには,細胞骨格のようなタンパク質と,それから形成される細胞内繊維構造をもっていたりして,この点でも真核生物に近い性質です.
クレン古細菌という可能性
クリックして拡大
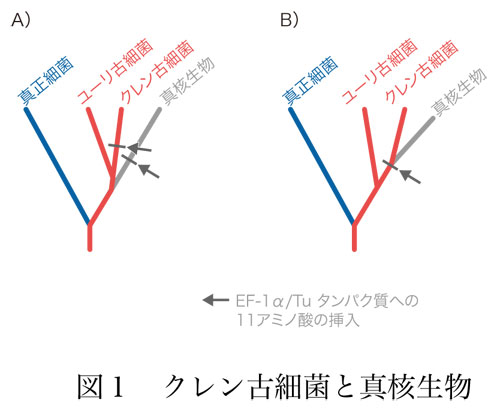
古細菌はユーリ古細菌とクレン古細菌の2つに大別されます.ユーリ古細菌の方がメジャーなグループで,メタン細菌や高度好塩菌のほか,サーモプラズマもここに含まれます.クレン古細菌には超好熱菌が含まれます.ちょっと驚くべきことですが,クレン古細菌と真核生物とが1つのグループであるという,エオサイト仮説があります(クレン古細菌の別名をエオサイトといいます).
1992年に,タンパク質合成系で働くEF-1α/Tuというタンパク質の中のGTPを結合する部位についてアミノ酸配列を調べると,クレン古細菌と真核生物とで共通に約11個のアミノ酸の挿入配列があるというエオサイト仮説を支持する結果が出ました.ユーリ古細菌にも真正細菌にもこの挿入配列がありません.挿入はクレン古細菌と真核生物に分かれた後で,それぞれ独立に同じ場所に同じ配列の挿入が起きたと考えるか(図1A),クレン古細菌と真核生物の共通祖先の段階で起きたと考えるか(図1B)といえば,後者を採るのが自然かつ単純で妥当でしょう.2008年のアメリカ科学アカデミー紀要の論文では,53個の遺伝子を対象とした解析から,クレン古細菌と真核生物の共通祖先の分岐を支持すると報告しています.
2.真核生物はDNAを貯蔵する核をもった
核をもつのが真核生物である
クリックして拡大
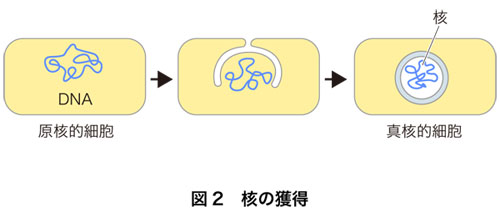
真核生物の細胞は,二重の膜で覆われた核をもった.これが,真核生物が後に大きく展開する第一歩になった重要なできごとであると思います.核の有無は,真核生物と原核生物とを区別する基本的な違いです.核は二重の膜で被われているので,図2のようにできたものと想像されています.遺伝子(DNA)を収納する特別なコンパートメントをもてたことで,DNAを大量に安定に保持できるようになりました.
核には高分子が出入りするしくみが必要
ただ,DNAを鋳型にして作られるRNAを核から細胞質へ輸送したり,核内で働く酵素やクロマチンタンパク質など,さまざまなタンパク質を細胞質から核へ輸送するには,核膜孔という特別な構造を用意することが必要でした.細胞質から核へ,あるいは核から細胞質へと行き来して機能するタンパク質は非常にたくさんあります.文章にすると簡単ですが,実際にはかなり複雑な機構が必要です.
真核生物のDNA量は多い
まず,DNAの量は,現在の原核生物と真核生物の間で大きな違いがあります.大腸菌とヒトを比べたとき,概略的にはヒトDNAは大腸菌の約1,300倍あります.ヒト体細胞1つは,約2mという長さのDNAをもちますが,これを50万倍に拡大すると,直径1mmで長さ20kmの糸状分子が46本存在する(あわせて約1,000km)ことになります.偶然かも知れませんが,細胞の体積も大体1,000倍違います.真核生物は細胞が大きくなり,DNAを保持する専門の核というコンパートメントをもったことで,DNA量を増やしても生きられるようになったことが,第6回で説明するように,真核生物が画期的な展開を果たす大きな出発点であったと思います.
真核生物の遺伝子数は意外に少ない
クリックして拡大
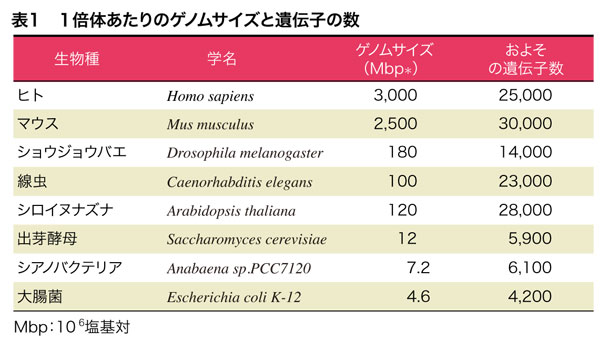
DNA量には大きな違いがあるものの,遺伝子の数は,大腸菌で4,300個,ヒトで25,000個といわれています(表1).大腸菌に比べてヒトはわずか6倍程度でしかありません.ここでいう遺伝子とは,大部分はタンパク質の構造を決めている遺伝子です.タンパク質の構造情報は,DNAの塩基配列からmRNAの塩基配列として転写されて,タンパク質が合成(塩基配列からアミノ酸配列への情報の翻訳)されます.真核生物では,選択的スプライシングによって,タンパク質としては10万種類くらいできると考えられていますが,それでも大腸菌の24倍でしかありません.バクテリアとヒトの複雑さの違いを考えると,遺伝子数の違いは信じられないほど小さいと思います.
これは,真核生物では,遺伝子の使い方にかなりの工夫があることを想像させます.真核生物は,限られた数の遺伝子をうまく使って,多様な生き物を作っているらしい.
遺伝子以外のDNAをもっている
タンパク質の構造を決める遺伝子の部分は,大腸菌ではDNA全体の90%以上ですが,ヒトでは1.2%程度でしかありません.イントロンや遺伝子発現調節領域など,遺伝子に関係するDNA全部を合わせても約25%です.残りの75%は,多くの繰り返し配列をもつことなどの特徴がありますが,機能的な必要性は不明です.ただ,個体が生きていくためのレベルでは無駄な存在に思える豊富な繰り返し配列とか,イントロンをもつことが,進化の過程における長い目でみると,遺伝子の多様化に強力な力として働き,さらに遺伝子の発現調節の複雑なしくみを可能にしました(第6回で述べます).
真核生物は複数の直鎖状DNAをもった
原核生物の多くは環状二本鎖DNAをもっており,DNA末端がありません.これに対してすべての真核生物の核内DNAは,直鎖二本鎖DNAをもっています.大量のDNAを,何本もの直鎖二本鎖DNAに分けて核内に収めています.真核生物のDNAが直鎖状であることが好都合,あるいはそれが必要であるという理由は,わかりません.
末端の始末
直鎖状DNAの末端部分に,テロメアという特殊な塩基配列をもった部分があります.短い塩基配列の繰り返しです.ヒトを含めた哺乳類では,5′-GTTAGG-3′という6つの塩基配列を単位として,数千~数万回繰り返した構造です.この部分には遺伝子はありません.テロメアDNAは末端の一本鎖(Gテイル)を使ってループを巻いていて,これにテロメア配列に結合するタンパク質が結合して,DNA末端を隠すのが役割と考えられます.末端の存在はDNA切断の非常事態と認識され,直ちに結合酵素が働いてしまうので,隠しておく必要があるわけです.
テロメアは複製できない
DNA複製機構のもつ性質から,直鎖状のDNAではDNA複製が起きるたびに,新しく作られるDNA鎖は必ず末端が短くなります.細胞が増えるためには,その前に必ずDNA複製が必要なので,細胞が増えるたびにDNAが短くなることは困った問題です.環状DNAではこういうことは起きません.真核生物の細胞にはテロメラーゼという酵素があって,複製の度に短くなるテロメア配列を延長して,長さを戻します.直鎖二本鎖DNAをもっている真核生物のDNAを安定に保持するためには,テロメアとテロメラーゼという特別な工夫が必要なのです.
テロメラーゼは最初の逆転写酵素か
テロメラーゼという酵素は,酵素タンパク質と鋳型RNAの複合体で,鋳型RNAを使って1本鎖DNA(Gテイル)を延長合成する逆転写酵素です.Gテイルが十分長くなれば,それを鋳型にして,通常の複製酵素によって相補鎖が合成されると考えられます.真核生物にみられるレトロウイルスやレトロトランスポゾンにはいずれも逆転写酵素が働いており,構造的な共通性をもったファミリータンパク質です.真核生物が直鎖状DNAをもったときにテロメラーゼが同時に用意されたものとすれば,真核生物における一番古い逆転写酵素です.
3.真核生物はクロマチン構造をもった
クロマチン構造
真核生物のDNAは,ヒストンという強い塩基性のタンパク質と強固な複合体を形成して,クロマチンという構造を作っています.4種類のヒストンが2つずつ集合した8量体を形成し,これにDNAが巻き付いています.真正細菌でも,DNAは裸で浮かんでいるわけではなくタンパク質と複合体を作っていますが,ヒストンとは全く別のタンパク質です.古細菌の場合は,DNAはヒストンに似たタンパク質と結合して,クロマチンのような複合体を作っているものが多くあります.真核生物ができたとたんに無から有を生じたわけではなく,こういう基本的なことについて,古細菌から真核生物への進化的なつながりがあるわけです.
塩基性タンパク質としっかり結合して,しかも高次のらせん構造を作ることで,非常に細くて長いDNAの糸を,比較的太くて短い糸(クロマチン)に仕立て上げることで,大切な遺伝子を安定に保存することに大いに役立ったと思います.クロマチンはDNAの安定な保持には好都合でしたが,DNAの複製や転写はちょっと面倒になりました.複製や転写の際には,その部分でタンパク質を外したり,タンパク質との結合を緩めることが必要だからです.多細胞生物ではむしろこのことを積極的に利用して,異なった種類の遺伝子を異なった組織や臓器で発現させる,新たな調節機構として展開するに至りました.
核骨格とクロマチンのコンパートメント化
核膜の内側と核の内部には,繊維性の核骨格という構造体があり,これにクロマチンが一定の規則性をもって結合しています.長い毛糸のようなクロマチンを核内にきちんと収めるためにも,細胞分裂時にクロマチンから染色体にまとめあげるためにも,DNAの複製や転写を実行する場としても,発現する遺伝子領域と発現することのない遺伝子の領域を区別して収納するためにも,核骨格の存在と,それによるクロマチンの核内への規則的な収納(コンパートメント化)機構が働いています.DNAの量的増大にはこういうシステムの並行的な成立が不可欠だった.
細胞分裂時には染色体を形成する
細胞が分裂する際には,それぞれのDNAからなるクロマチン糸はさらに凝縮して,太くて短い染色体になります.このとき,通常は核膜構造が消失します.ヒトの体細胞の場合,染色体は46本できますが,1本の染色体は複製を終えたばかりの2本の染色分体からできています.92本もある長い糸をもつれたり切れたりしないように実行するわけです.
具体的にはセントロメアDNAという特別な繰り返し配列とセントロメアに結合するタンパク質によって形成される,それぞれの染色分体の動原体に結びつく紡錘糸などが働く複雑なプロセスが進行します.そして,それぞれの染色分体を2つの娘細胞に誤りなく分配します.
プロセスのチェック機構
また,このプロセスのあちこちで,そこまでのプロセスが正しく進行しているかどうかをチェックし,正しくなければその先へ進ませない,というチェックポイント機構(チェック・アンド・ゴー機構)が働いています.チェックがOKになると先へ進行しますが,その場合,必要タンパク質の分解が起きるなど,後戻りできないように仕組まれています(ラチェット機構).そのあと,核膜が再生して核が復活し,染色体はクロマチン糸にほぐれ,細胞質が2つに分かれます.真核生物が遺伝情報を正しく2つに分けるには,このような有糸分裂の機構の獲得が必要でした.
次回は,真核細胞がどのように進化してきたのか,オルガネラはどうやってできたのか,最新の知見をご紹介します.現在進行形でオルガネラ化しているのかもしれない真性細菌もいるようです.・・・続きは次回!

プロフィール

おすすめ書籍


















