![第5回 真核生物の誕生2 1.真核生物は複雑な細胞内構造をもった [分子生物学講義Web中継~生物の多様性と進化の驚異]](images/banner03.gif)
- [SHARE]
- ツイート
")

第5回 真核生物の誕生2
- 真核細胞に進化するために重要な機能は「貪食」だった?
- アブラムシは新しいオルガネラを獲得中?
- ・・・など,驚きの視点が満載.
1.真核生物は複雑な細胞内構造をもった
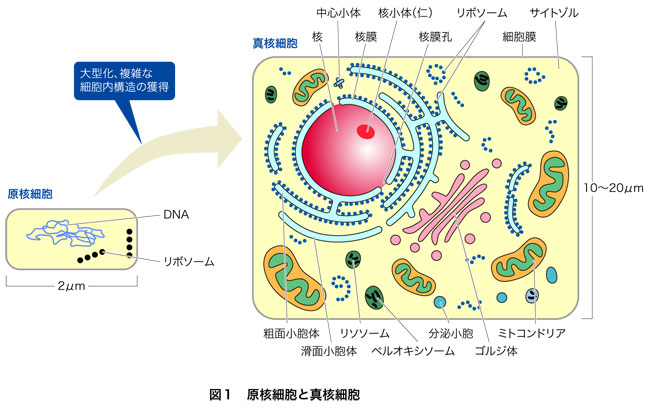
大型化した真核生物は大きな核と大きくて複雑な細胞質をもつ
クリックして拡大
真核生物は核をもってたくさんのDNAをもてるようになり,細胞質も大きくなりました.大きいだけでなく,原核生物との違いとして特徴的なのは,細胞質にさまざまな種類の細胞内小器官(オルガネラ)がぎっしり詰まっていることです(図1).オルガネラは,膜構造で囲まれた構造体で,さまざまな機能を分担しています.誕生したばかりの古細菌の細胞膜はテトラエーテル型リン脂質でしたが,真核生物はどこかの時点で環境温度の低下に見合ったエステル型リン脂質の細胞膜に置き換えて,それが現在まで続いています.
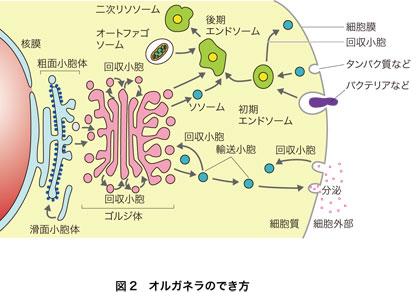
オルガネラのでき方と相互の関係
オルガネラは互いに関係があります.図2の下の方に滑面小胞体がありますが,ここで細胞質から脂質が膜に組み込まれて脂質膜が拡大します.これにリボソームが結合すると粗面小胞体になり,ここで合成されるタンパク質には,膜タンパク質として膜に組み込まれるものと,小胞体内部に蓄えられるものがあります.
クリックして拡大
粗面小胞体から輸送小胞が出芽してゴルジ体へ移動して融合し,ゴルジ体で膜や脂質に糖鎖の付加という修飾が起きます.ゴルジ体から,リソソーム独自の膜タンパク質や内部に分解酵素類を濃縮した小胞が出芽して,リソソームになります.リソソームは多種類の分解酵素をもった袋で,細胞外から取り込んだ高分子や固形物などの初期エンドソームや,古くなったオルガネラなどを取り囲んだファゴソームと融合して,後期エンドソームになって内容物を消化します.
他方,ゴルジ体からは,細胞膜や分泌する物質を含んだ小胞が出芽し,細胞膜の方向へ運ばれてやがて細胞膜と融合し,細胞膜を供給したり,内容物を細胞外へ分泌したりします.輸送体としてのたくさんの小胞は先方のオルガネラと融合しますが,内容物を先方へ渡した後,回収小胞として出芽して元の場所に戻るといった芸の細かいことが行われています.
膜トラフィック
クリックして拡大
このように,オルガネラ全体として互いに関係しており,膜の移動という意味でこのような動きを膜トラフィックといいます.膜だけでなく,膜で包まれた内容物も移動します.真核生物の細胞が大きく複雑になることができたのは,単なる拡散に頼ることなく,膜トラフィックによって積極的に物質を移動させる機能を獲得したからであるともいえます.現在の動物細胞ではこのようなトラフィックが稼働していますが,図3のような単純なところから,このような複雑な系がどのように成立したかはよくわかっていません.
2.真核生物は細胞骨格をもった
サイトゾル中の構造物
オルガネラの間を埋める無構造のサイトゾルは一見無構造にみえますが,案外多くの構造物があります.繊維性の細胞骨格のほか,タンパク質合成の場であるポリソーム(リボソームがmRNAでつながったもの)があります.プロテアソームという巨大な分解酵素複合体もあります.これは64個ものタンパク質が集合した樽のような形をしていて,樽の蓋の部分で分解すべきタンパク質とそうでないタンパク質を識別して,分解すべきタンパク質を引き入れて,内部を向いて働く複数のタンパク質分解酵素が消化します.サイトゾルにはこのほか,解糖系の酵素をはじめとするさまざまな代謝系があり,また,細胞膜から細胞質内や核内へ,あるいはその逆の経路でさまざまな信号を伝達するシグナル伝達系のタンパク質や酵素などが,緩やかな一定の構造をもって配置されているものと考えられます.
細胞骨格
真核生物は,細胞内に細胞骨格という繊維状の構造をもっています.オルガネラは膜で囲まれた構造物を指すので,細胞骨格はオルガネラには含めません.細胞骨格には主に3種類あって,ミオシンと共同して細胞運動を司るアクチン繊維(アクチン),キネシンやダイニンと共同してタンパク質・オルガネラ・小胞の細胞内移動を司る微小管(チュブリン),細胞の丈夫さを司る中間径繊維(ケラチン,ビメンチンなど)です.
細胞極性の成立と維持
上皮細胞は,極性をもっています.極性というのは方向性のことです.例えば腸の上皮なら,消化酵素を外部へ向かって分泌する一方で,栄養物を外部から体内に向かって吸収するという方向性をもっています.自由端面(頭頂部)の細胞膜と,側方と底面(側底部)の細胞膜とでは,輸送タンパク質の分布が異なるわけです.頭頂部では栄養素を細胞外から細胞内へ輸送し,側底部では同じ栄養素を細胞内から細胞外へ輸送しなければなりません.これができるためには,輸送タンパク質の種類によって,細胞膜への別の部位まで運ぶことが必要です.
上皮細胞では構造的にも極性があります.細胞の1つの面は自由端ですが,側面は隣の細胞とさまざまな接着構造によって接着し,底面は基底膜という細胞外の構造体にしっかり接着します.接着タンパク質の細胞膜における分布に極性があるわけです.構造的にも機能的にも極性があるわけですが,極性構造の構築にも,極性をもった機能を維持するにも,接着タンパク質と細胞骨格とモータータンパク質が協調して働いています.これは,多細胞動物が組織を構築し,器官を構築して,適切な構造と機能を保つために必要な基本的な機能の1つです.
貪食という機能
白血球が這い回ってバクテリアを貪食するという話は聞いたことがあるでしょう.原生生物のアメーバが他の細胞を餌として取り込むのも貪食です.これらの細胞は顕著な例ですが,ほとんどの細胞がこの機能をもっています.細胞骨格を手に入れた真核生物は,運動性と貪食性を獲得したことで,餌の確保が画期的に有利になりました.積極的にえさを探しに出歩けて,餌をみつけて高分子でも固形物でも貪食し,貪食したものを細胞内で消化できます.運動して到達できる周囲に餌がある限り,生きのびられるようになった.これで動物型生物の原型ができた,ともいえます.これは,従属栄養生物にとって非常に大きな進歩であったと思います.
共生も貪食の結果かもしれない
もう1つ重要なことは,細胞内共生には貪食が働いていた可能性です.好気性細菌を貪食したとき,大部分は消化して餌になったでしょうが,一部は生きのびて共生状態に入った.それでミトコンドリアができた.葉緑体も同様です.貪食がそういう役割を果たしたとすれば,真核生物の進化にとって画期的に重要なことです.
運動性と貪食性を獲得する前提として重要なことは,真核細胞が硬い細胞壁を失ったことです.細胞壁があるままでは運動性も貪食性も発揮できない.真核生物の誕生は細胞壁をもたない古細菌からなのか,真核細胞になった後で細胞壁を失ったのかは不明です.現在の原生生物の中にも二次的に堅い殻をもつものがありますが,殻のあちこちに穴が空いていてそこから細胞質を伸ばして運動するような例はあり,丈夫さを保ちつつ運動性も発揮して,栄養素のあるところを捜して歩く,といった途中プロセスがあり得ます.想像に過ぎませんが,そのうち,そういう微化石がみつかる可能性だってないわけではない.
進化的な連続性
細胞骨格は真核生物にしかなく,原核生物にはない,といわれてきました.無から有が生じたのだろうか.つい最近,バクテリアにも,アクチンやチュブリン,中間径繊維と似た細胞骨格様のタンパク質があり,それからできた繊維性構造が細胞内にあること,細胞内の物質や構築物の移動に働いているなど,真核生物と類似していることがわかりました.原核生物のアクチン様タンパク質はATPと結合するとか,チュブリン様タンパク質はGTPと結合するなどの性質にも,真核生物のアクチンやチュブリンとの共通性があります.いきなり無から有を生じたわけではなく,ちょっとした工夫とやりくりが進歩をもたらした可能性が高いのです.なぜ最近までわからなかったのだろうと不思議に思うでしょうが,その気で調べなければ,見るもの見えずということはいくらでもあるのです.マイコプラズマでは,真核生物にはみられない細胞骨格と運動装置をもっていることも,最近わかりました.バクテリアの類だって,それなりに工夫しているわけです.
3.真正細菌の共生とオルガネラ化
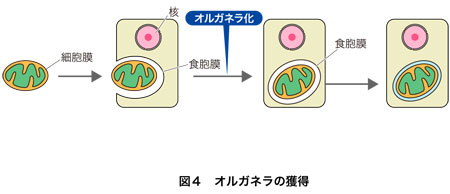
クリックして拡大
ミトコンドリアも葉緑体も,かつて共生した真正細菌の名残であることがわかっています(図4).
好気性真正細菌の細胞内共生
およそ20億年前に酸素濃度が現在の濃度の1%を超え,好気的酸化が可能な環境になるとすぐに,真正細菌のなかから好気性バクテリアが誕生し,好気性バクテリアが誕生すると間もなく真核細胞内に共生をはじめたと考えられます.遺伝子構造の共通性からみて,共生したバクテリアは,現在の真正細菌のなかのαプロテオバクテリアというグループの,リケッチアに近い好気性細菌と考えられます.ただ,ほとんど無酸素状態の深海底にいた可能性のある古細菌と,海面近くの酸素濃度が高いところに生息していたであろう好気性バクテリアが,どのように出会ったかには問題があります.現在のクレン古細菌のなかには,比較的低温で生育するものや,好気性のものさえあるので,こういうタイプのものが古くからいれば,出会うチャンスはあったかも知れません.
ミトコンドリアの成立
共生した好気性バクテリアは,独立した細胞としてのさまざまな機能を消失して単純化し,やがてミトコンドリアになりました.取り出したミトコンドリアは,単独で生きていくことができなくなっています.こうして,古細菌に由来する細胞質がもっていた,嫌気的に有機物を部分分解する代謝経路と併せて,ミトコンドリアで酸素を使って有機物を最終的に酸化し,効率よくエネルギーを生産して,エネルギー貯蔵分子であるATPを合成する機能を身につけました.真核生物は好気性生物として,莫大なエネルギーを生産・消費できるようになり,活発な活動をすることができるようになりました.たくさんのミトコンドリアを保持するには,細胞質が大きくなり,かつ,酸素濃度が上昇して酸素供給が十分になることが必然でした.酸素濃度の上昇,シアノバクテリアの共生,大型真核生物の誕生が,およそ20億年前に平行して起きたことが理解できます.
ミトコンドリア遺伝子の核への移行
好気性バクテリアが真核生物の細胞質に共生したとき,単独で生活するのに必要な遺伝子の多くを消失しました.不思議なことにミトコンドリアでは,ミトコンドリアの形成に必要なたくさんのタンパク質の遺伝子は核へ移行して,核内遺伝子として存在しています.
ミトコンドリア遺伝子を核へ移行させた方がよい理由と移行したしくみについてはよくわかっていません.動物のミトコンドリアのゲノムは20kb以下と小さく,含まれる遺伝子数も50個以下と少ないのが普通ですが,植物では大きな幅があり,ゲノムサイズで500~2,500kbpにもおよぶものがあるといわれます.植物ミトコンドリアゲノムには,葉緑体ゲノムから移動したものが含まれる場合があるといわれます.なお,葉緑体の場合にも,かなりの遺伝子が核に移行しています.
リケッチアは今でもミトコンドリアを後追い
遺伝子解析から,ミトコンドリアは真正細菌のリケッチアに一番近いといわれます.現在のリケッチアはすべてが寄生性で,発疹チフスやツツガムシ病などの病原菌の仲間ですが,動物だけでなく植物にも寄生します.植物のこぶ(クラウンゴール)を作るアグロバクテリウムや窒素固定で有名な根粒菌もこの仲間です.宿主の細胞内で増殖し,細胞外で増えることはできません.ゲノムサイズは真正細菌のなかでは小さく,1,100kbp程度のものです.代謝的には宿主細胞に依存しているので,代謝系遺伝子のほとんどを失っていますが,クエン酸回路や電子伝達系を保持しATP合成を行うところはミトコンドリアと似ています.ミトコンドリアの後を追って,単純化への道を歩んでいるようにみえます.ミトコンドリアとの違いは,ノミ,シラミ,ダニ,ツツガムシなどを介して感染することと,感染した宿主に病気を起こすことです.
コラム:オルガネラ化に向けて現在進行形(?)の真性細菌
原核生物と真核生物との共生関係は現在でも非常にたくさんの例があります.オルガネラといえるくらいまで進んでいるものもあります.多くのなかから2つだけ紹介しておきます.
アブラムシが主食とする植物の篩管液にはグルタミンとアスパラギン以外の必須アミノ酸が含まれておらず,アブラムシ自身の代謝系では必須アミノ酸を合成できないので単独では生きていけません.しかし,ブフネラという真正細菌が細胞内に共生していて,必須アミノ酸を合成して供給してくれるので,アブラムシは生きていけます.ブフネラは単独に生きるために必要な遺伝子の多くを失っているために,取り出して単独で生きていくことはできません.ブフネラはアブラムシの卵子から子へ伝えられるという点でも,オルガネラに近い存在といえます.ただ,ブフネラはアブラムシの全細胞に存在するわけではないので,オルガネラとはいわれません.この共生関係は2億年以上も続いているといわれます.
節足動物(昆虫,クモ,ダンゴムシその他)や線虫などに広く寄生している,ボルバキアというリケッチアの仲間の真正細菌がいます.さまざまな器官に感染しますが,なかでも精巣や卵巣に感染して生殖能力に大きな影響を与えます.感染した雄は死んだり,雌化したりします.感染した雌では単為生殖します.卵子を通じて子孫に伝わりますが,成熟した精子には存在できないために精子から子孫には伝わりません.オルガネラ化してはいませんが,卵子を通じて子孫に伝わるところや,自身の遺伝子の一部を宿主細胞に移行させることはオルガネラ的です.個体間での感染が起き,種を超えた個体間で感染することもあります.生きる工夫を言い出すと切りがありませんが,ボルバキアには持続感染しているウイルスがいて,種を超えて感染した際にウイルスが活性化して,ボルバキアが新しい宿主に住みやすくなるように遺伝子変異を促進するといった複雑なこともあるらしい.
35億年の歴史をもつ原核生物はついに多細胞生物にはなりませんでしたが,真核生物はやがて多細胞生物を生み出します.多細胞動物の誕生の先にヒトの誕生もあるわけですが,多細胞動物誕生のために何が必要だったのか,第6回で少し詳しく考えてみます.多細胞化するために必要な準備は,単細胞のうちになされたと考えられます.
次回は,真核細胞が,ヒトを含めた真核多細胞生物になるまで,どのようなことが必要だったのか,最新の知見をご紹介します.原核細胞が多細胞化への道を進まなかったなかで,真核細胞はいろいろと複雑な準備をしていたようです.・・・続きは次回!

プロフィール

おすすめ書籍


















