![第6回 生命の多細胞化に必要だったこと 1.ラクシャリー遺伝子というもの [分子生物学講義Web中継~生物の多様性と進化の驚異]](images/banner03.gif)
- [SHARE]
- ツイート
")

第6回 生命の多細胞化に必要だったこと
- 1つの遺伝子が異なる生物でも機能する?
- ラクシャリー遺伝子はハウスキーピング遺伝子から誕生した!
- ・・・など,驚きの視点が満載.
1.ラクシャリー遺伝子というもの
多細胞生物の特徴
単細胞から多細胞への変化は,細胞の誕生,真核細胞の誕生に次ぐ,進化の上で第3の画期的なできごとであったと思います.多細胞化は単細胞では限界のあった,複雑な構造と機能をもてるようになり,生物としての多様な展開を可能にしました.また,多細胞生物というのは,構成細胞1つ1つが機能的にも形態的にも分化し,役割り分担していて,細胞集団全体(個体)として一定の形態的特徴をもち,個体としての機能的な統合がある,という特徴をもっています.単純にいえば,脳を作るには脳の遺伝子がいる,心臓を作るには心臓の遺伝子がいる,できた脳や心臓の働きを維持・調整するにもそれなりの遺伝子がいります.そういう遺伝子,ラクシャリー遺伝子は,単細胞のバクテリアには必要がなかったものです.ラクシャリー遺伝子を用意しなければ,多細胞化は実現しなかったと考えられます.第6回では,動物の多細胞化に必要な遺伝子をどのように用意したかについて述べることにします.
進化を進める遺伝子の変化
たくさんのラクシャリー遺伝子を準備したのは,真核生物特有のしくみの獲得によります.その前提として,細胞が格段に大きくなったこと,核というコンパートメントができたことで,たくさんの量のDNAを安定に保持できるようになったことが,すべての出発点であったと思います.遺伝子を増やす方法をまとめて紹介します.
2.遺伝子を増やす方法
遺伝子セットの倍数化
クリックして拡大
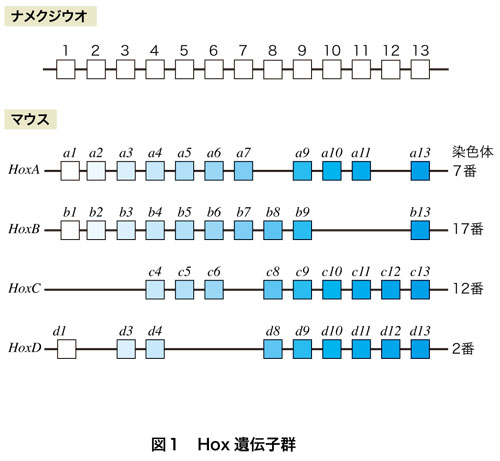
細胞あたりのDNAの量をドーンと増やす方法があります.通常の細胞は2倍体で,遺伝子を2セットもっているわけですが,比較的容易に4倍体の細胞になり,4倍体の個体になることがある.4倍体の親同士からは,ちゃんと4倍体の子孫が維持できる.魚類や両生類には今でもしばしばみられることです.動物の発生過程で形作りに働く重要な遺伝子として,Hox遺伝子群がありますが,これが倍数化の好例です.動物の体の頭からしっぽにかけて,発生過程で体の各部分の特徴的な構造を作り出すことに働く13個の遺伝子群が,まとまって配列しています(図1).脊索動物のナメクジウオでは半数体あたり1セット,2倍体細胞としては2セットあります.これが,無顎類のヤツメウナギでは半数体当たり2セットに増え,魚類,両生類,爬虫類,哺乳類では半数体当たり4セットと増えます.ゲノムレベルでの倍化が2回にわたって起きた結果,哺乳類では4倍になっているわけです.Hox遺伝子部分だけが増えたのではなく,染色体1本のレベルで倍加した証拠に,Hox遺伝子だけでなく,その染色体に乗っていた他の遺伝子も倍化していることがわかっています.増えたHox遺伝子群はそれぞれ変異して機能を失ったり,機能分担をするようになったので,哺乳類の4セットは,互いによく似ているけれども完全に同じ遺伝子のセットとして存在しているわけではありません.脊索動物から脊椎動物へ,脊椎動物の無顎類から顎口類へ進化するあたりで起きたゲノムの大規模な増幅は,さまざまなラクシャリー遺伝子の劇的な増加とも関係していると考えられています.
有性生殖による遺伝子組換え
減数分裂の過程でのDNAの組換えは,減数分裂の過程を光学顕微鏡で観察していた時代から,染色体交叉として知られていたものです.ヒトの場合,1回の減数分裂あたり,およそのところですが,染色体1本に1回の組換えが起きる.母親由来の1番DNAと父親由来の1番DNAの間で組換えを起こすと,母親の配列と父親の配列をもってつながった1番DNAが,2本できます.母親と父親の塩基配列をモザイク状態に保持したDNAが2本できるわけです.組換えの起きる場所はランダムだから,生殖細胞の遺伝子の多様性はほとんど無限大である.
減数分裂の際には,積極的に組換えを起こして,遺伝子を積極的に多様化させていると思われる理由が少なくとも2つあります.1つは,相同染色体の対合というプロセスがあることです.減数分裂が,2倍体の細胞から1倍体の生殖細胞を作ることだけを目的とするなら,母親由来の染色体と父親由来の染色体とを対合させる必要性は全くありません.
もう1つは,異常に高いDNAの組換えの頻度です.組換えは,体細胞でも起きなくはありませんが,減数分裂の際に比べてせいぜい1万分の1以下です.ところが,減数分裂の場では,DNAを切って繋ぎ変える,組換え酵素があらかじめ集合しています.これらを考えると,減数分裂とは,積極的に組換えを起こす場として仕組まれているようにみえます.
遺伝子組換えによる遺伝子重複
遺伝子組換えが2本のDNAのずれた場所に起きると,1本のDNA上には同じ遺伝子が2つ,他方のDNA上にはゼロになってしまうことがあります.同じ遺伝子を2つもったDNAでは,遺伝子の重複が起きたことになります.真核生物にはこのようにしてできた遺伝子ファミリーがたくさんあり,それぞれが少しずつ変異を重ねて機能を分担しています.
エキソンシャフリングによる新しい遺伝子の構築
クリックして拡大
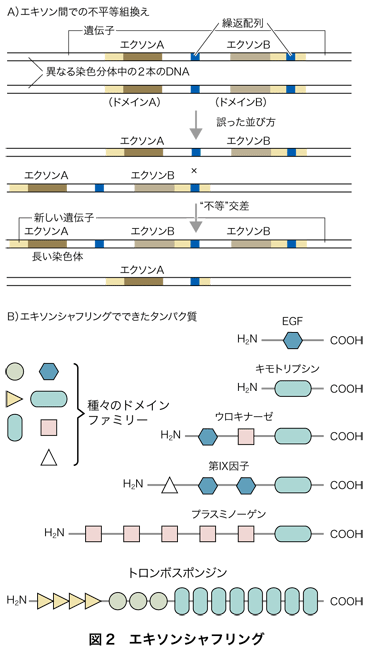
トランプの札を混ぜ合わせる(ランダム配列化する)ことをシャフリングといいます.減数分裂の際に,イントロン部分でDNA組換えが起きることによってエキソンを混ぜ合わせることを,エキソンシャフリングといいます.機構的には遺伝子重複と同じことですが,組換えが遺伝子の間ではなく,遺伝子内部のイントロンの間で起こります.繰り返し配列がイントロン中にしばしばみられ,ここがDNAの相同組換えに使われて,エキソンがシャッフルされるわけです(図2).それぞれのエキソンが,タンパク質の構造的・機能的な単位構造(ドメイン)を構成する場合がしばしばみられ,エキソンを組合わせることは,構造的・機能的単位を組合わせることである,といえます.
クリックして拡大
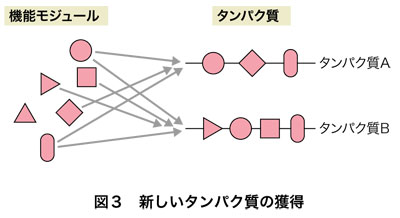
エキソンシャフリングは,新しい構造をもった遺伝子を作り出し,その遺伝子情報から新しいタンパク質を作り出す画期的な方法の提示でした.エキソンというすでに機能をもっている既存の単位(ドメインあるいはモジュール)を無数に組合わせ,そこから,新しい機能をもったタンパク質の遺伝子ができる可能性が示されたわけです(図3).
遺伝子の水平移動とトランスポゾン
遺伝子の水平移動もラクシャリー遺伝子の準備に貢献した可能性があります.大昔,細胞が誕生して古細菌から真正細菌や真核細胞が分かれるまでの間,DNAの水平移動が頻繁にあった可能性を第3回で紹介しました.バクテリアがDNAを取り込む形質転換や,動物細胞がDNAを取り込むトランスフェクションも水平移動の応用といえ,研究に汎用されています.
トランスポゾンといって,細胞DNAから抜け出し,細胞DNAのあちこちに入り込む,細胞内の寄生虫のような小さなDNAもあります.DNA型トランスポゾンやレトロトランスポゾンなど,いくつかの種類があります.
3.増えた遺伝子をやりくり
増やした遺伝子をやりくりする
単細胞のときには1つしかなかった遺伝子が,やがて重複やエキソンシャフリングを繰り返し,それぞれが少しずつ変化してファミリーを形成し,機能的に多様化する.こうして新しい遺伝子ができ,新しいタンパク質が作られ,有害でなければ排除されることもなく,種の集団のなかではさまざまな変異遺伝子が温存される.そうやって増えて多様化した遺伝子が蓄積していることで,あるとき,それに加えてたった1つの遺伝子の変化が起きると,それまでは有効な働き場がなかったタンパク質をやりくりして,結果的に新しい機能を誕生させることはありうることです.
眼をもたなかった動物に眼ができる,脊索をもたなかった動物に脊索ができるといった結果を生じる,などという大げさなことは本当に稀で極端な例でしょうが,当面は役に立たないようなたくさんの遺伝子を蓄積することは,大きな変化への準備段階として有効です.生き物は,これらの遺伝子を特に利用することなく保存している場合もあれば,やりくりしながら使っている場合もある.生き物というものは,やりくりの天才でもあるのです.
遺伝子のやりくり構築の例
脊椎動物はよく発達した目をもっていますが,目のレンズはクリスタリンというタンパク質が集合したもので,極めて透明性の高いものです.クリスタリンも多くのメンバーからなるファミリーで,α-,β-,γ-クリスタリンは脊椎動物全部に共通です.驚いたことに,これらはいずれも,解糖系のエノラーゼや乳酸脱水素酵素,尿素回路のアルギノコハク酸リアーゼの他,プロスタグランジンF合成酵素と構造的に似ていることがわかりました.構造的に似てはいても,多くは酵素としての活性をもつわけではありません.ただ,εクリスタリンについては実際に乳酸脱水素酵素活性ももっているといわれています.脊椎動物だけでなく,頭足類(イカやタコ)ではグルタチオン-S-トランスフェラーゼという酵素が,活性をもったままクリスタリンになっているといわれます.
副業(内職)タンパク質
異なる2つ(以上)の機能をもつタンパク質を,moonlight proteinと称します.ここで使うmoonlight は,昼間の仕事とは別にする『夜の副業』のことです.内職・夜なべ仕事といった感覚です.moonlight proteinは,性質の異なる2つの仕事(機能)をもったタンパク質のことで,こういうタンパク質は最近たくさんみつかっており,例えば極端な例ですが,グリセルアルデヒド-3-リン酸脱水素酵素(GAPDH)は,解糖系の酵素としての活性のほか,DNA修復時やDNA複製時のタンパク質複合体に含まれて働き,男性ホルモン受容体タンパク質が遺伝子DNAに結合して転写促進する際の促進タンパク質としても働き,tRNAの輸送にも働き,細胞死(アポトーシス)のプロセスでも役割を果たし,エンドサイトーシス(貪食)の際や細胞内の小胞輸送にも微小管の重合にも働くのだそうです.2つどころか山ほど副業をしているらしい,というか,ここまでくるとどれが本業なのかわからない.
ハウスキーピング遺伝子からラクシャリー遺伝子ができる
クリスタリンの場合,解糖系酵素のようにバクテリア時代から存在する非常に古い歴史をもつ酵素タンパク質から,遺伝子重複によって酵素遺伝子が増え,さらに遺伝子変異によってレンズタンパク質になった,というプロセスが考えられます.2つ以上の機能をもつタンパク質があったとき,どちらが主業でどちらが副業かは単純にはいえませんが,今まで知られた例ではクリスタリンに限らず,機能の1つは解糖系の酵素などであることが多いようです.解糖系酵素の遺伝子は,原核生物にも真核生物にも共通に存在するハウスキーピング遺伝子で,生物界で最も古い歴史をもつ代謝系と考えられるので,こちらが主業(古くから携わってきた仕事)だったと考えられます.
進化の過程で,ハウスキーピング遺伝子しかもっていなかった原核生物を出発にして,真核生物がどのようにしてラクシャリー遺伝子を獲得するにいたったかは,大きな謎でした.ラクシャリー遺伝子の誕生は,無から有を生じることだったようにみえるからです.無から有が生じることは滅多にないけれども,既存のものをちょっと変化させて別の役割をもたせることなら,十分に可能性のあることです.moonlight protein発見の重要な意義は,解糖系酵素というバリバリのハウスキーピング遺伝子から,レンズのクリスタリンというバリバリのラクシャリー遺伝子が,遺伝子重複と若干の変異によって誕生する可能性が現実にありそうなことと示したところにあります.
同じ遺伝子が異なる生物で異なる役割りを果たすというやりくり
脊索を作るBra遺伝子は脊索動物では脊索を作るのに働いていますが,同じ新口動物の棘皮動物や半索動物にあるだけでなく,旧口動物の環形動物(ミミズなど)にもあり,さらに原始的な刺胞動物(クラゲの仲間)にもあります.これらの動物では,脊索を作ることではなく別の役割りを果たしています.眼を作る遺伝子であるPax6は,哺乳類の発生の初期には神経管の形成に,発生が進むと眼の形成だけだけでなく顔面の形成にも,成体になってからはホルモン形成のα細胞の誘導にも関係するといいます.1つの遺伝子がさまざまな動物で,さまざまな場面で,さまざまな細胞で,さまざまな異なった働きをするようにみえるのは,当該タンパク質の遺伝子が生物によって少しずつ変化して,機能はほとんど同じでも,一連の反応経路のなかで新しい働き方をもったためと考えられます.これによっても生物は新しい応答性を創生することができ,新しい表現形を生み出す可能性があるわけです.これも既存遺伝子のやりくり,タンパク質機能のやりくりの1つといえます.
コラム:重複によってできた遺伝子ファミリー
配列がよく似ているけれども細部では異なるファミリー遺伝子は重複によってできたと考えられています.例としては,さまざまなものがあるのですが,単細胞のときからもっていたタンパク質という意味では,オプシンファミリーが好例です.さまざまな生物が光受容タンパク質としてオプシンファミリーをもちます.ファミリーはすべて,膜に埋め込まれたタンパク質で,光のエネルギーをつかつて機能を果たすことで共通しています.例えば,哺乳類などでは視覚を司ります.しかし,古細菌のもつバクテリオロドプシンは細胞膜にあって,光のエネルギーを使って水素イオンを輸送するイオンポンプとして働いています.生存にとって必須の機能(ハウスキーピング機能)を担っていたバクテリアロドプシンのようなタンパク質の遺伝子が,重複して少しずつ機能的な変化をすることで,やがて視覚にも利用されるようになった,という歴史を示しているのかも知れません.
これまで,現在の分類と,地球誕生から多細胞化への準備について,わかりやすくご紹介いただきました.しかし,「進化の試行錯誤」と「その過程で誕生した生き物」は,とてもここでは語り尽くすことができません.そこで,8月下旬発行の単行本「分子生物学講義中継シリーズ」の最新刊では,「生物の多様性と進化の驚異」を井出先生に大いに語っていただきました! ここで紹介できないことが残念なぐらい,緻密なイラストと図が満載です! 生き物が大好きな人に自信をもってお薦めですので,ぜひ手に取ってみてください.

プロフィール

おすすめ書籍


















